葉の形づくりに関わる新しい遺伝子を探して
(1)はじめに
高等動物が胚発生の段階で、器官形成を終えて外界に出てくるのに対し、高等植物の胚では子葉、胚軸、幼根のみが形成されています。それ以外の器官は発芽後に茎頂・根端にあるメリステムと呼ばれる未分化な細胞群から作られます。茎頂メリステムはドーム上の一部が隆起して器官原基となり、細胞分裂と伸長により葉や花、茎などの新たな器官が次々と形成されます。これは動物の発生システムとは大きく異なる点です。
植物が既に分化した細胞から完全な個体を再生できる能力(分化全能性と言います)を持つことも、このような発生メカニズムの違いが関与しているのかもしれません。芽と葉は個体を再生する際、最初に生じる器官であり、植物の発生メカニズムを理解する上で、葉の発生の分子機構を明らかにするのは重要なことであると考えられます。
(2)葉の形づくり
葉は皆さんも御存知のように扁平な特徴的な形をした器官で、茎頂と根端を結ぶ軸を主軸として、茎頂メリステムから隆起した後、3つの軸、基部先端部軸、向背軸、中央側方軸に沿って成長し、扁平な形状の葉身を形成します。葉の表側(向軸側)と裏側(背軸側)では、異なる性質を持ち、向軸側では光合成に適した細胞に、背軸側ではガス交換に適した細胞に分化しています。葉の形づくりの分子機構は、まだよくわかっていませんが、ここ数年で、多くの遺伝子が葉の形態形成に関与することが明らかとなってきました。
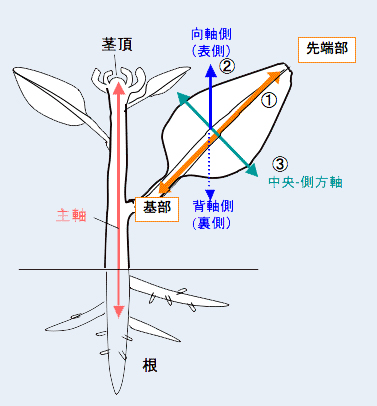
葉の表側(向軸側)と裏側(背軸側)では、それぞれに特異的な遺伝子群が発現しています。特異的な遺伝子は互いによく似ており、1つだけ壊れてもほとんど影響はありませんが、全てを壊す、あるいは表裏全体で強制的に働かせると、ラッパ状の葉や片方の性質のみをもつ棒状の器官が形成されます。最近は葉の分化にmicro RNAと呼ばれる小さなRNA分子が関わっていることを示唆するデータも報告されています。しかし、向軸側または背軸側に特異的な遺伝子群の発現を調節する機構はまだ十分解明されていません。双子葉植物に関しては、中心部の茎頂メリステムからの何らかのシグナルによる位置情報をもとに向背軸が決定されると考えられていますが、そのような因子はまだ見つかっていません。
また、茎頂で強く発現するclass I KNOXと呼ばれるホメオボックス遺伝子の葉原基での発現パターンが植物種によって異なっており、その発現パターンの多様性は葉の形態の多様性と関連があると考えられています。これらの遺伝子は、いずれも何かの遺伝子の転写を調節する転写因子として働くと予想されていますが、標的となる遺伝子は明らかになっていません。
このように多くの遺伝子が同定されつつも、その分子機構はまだ多くの謎が残っています。
(3)葉の形づくりに関わる遺伝子を探す
そこで、葉の形づくりに関わる新しい因子を見つけるために、葉の形が変化する変異体を見つけ、その原因遺伝子の同定を行っています。前述のように、葉の形態形成に関わる重要な遺伝子は、重複した機能をもつ相同な遺伝子が存在しているため、1つの遺伝子が壊れても形が変わらないことが多々あります。そこでシロイヌナズナのasymmetric leaves2 (as2)変異体を利用しました。

as2変異体は葉が左右非対称になる、葉の表面が凸凹になる、下向きにカールするなど、葉のさまざまな形態に影響を与えます。従ってas2変異体では葉の形態形成にかかわる別の遺伝子がもう一つ壊れるだけでも葉の形に影響が出やすくなると予想されます。実際に、as2に変異を起こさせた植物からは、葉の形が著しく変化した個体が多く得られました。その中には、AS2遺伝子が正常な状態では、葉の形にほとんど変化が見られないものもあります。得られた変異体を解析することで、植物の形づくりのメカニズムが少しでも解明されることを期待しています。