植物のDNA突然変異の抑制機構
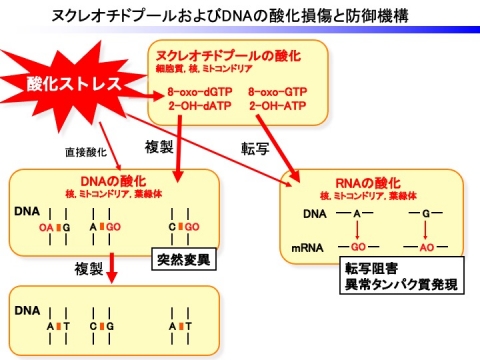
遺伝情報を担うDNAは生命活動が正常に営まれるためには安定に維持されなければならないが、DNAは内因性や外因性の要因で生じるAOSにより絶えず種々の損傷を受けています。DNAが酸化された場合、さまざまな損傷が生成し、DNAの突然変異を誘発するため、これらが修復されなければ、突然変異が蓄積し、細胞死や腫瘍化につながります。ヌクレオチドプール中のDNA合成の基質であるdGTPやdATPは酸化的ストレスにより8-oxo-dGTPや2-ヒドロキシアデニン(2-OH-dATP)となる。8-oxo-dGTPはシトシンと同程度の割合でアデニンと対合できるため、DNAの複製に際にG:C→T:AもしはA:T→C:Gトランスバージョン変異を引き起こします。
それに対して、生物は酸化ヌクレオチドの蓄積による細胞障害を抑制するために巧妙な防御系を備えています。すなわち、バクテリアや動物では、酸化ヌクレオチドである8-oxo-dGTPなどに対して加水分解活性を有するNudix hydrolaseによる酸化ヌクレオチド浄化機構や、酸化塩基除去修復機構が存在し、DNAが健全に保たれています。
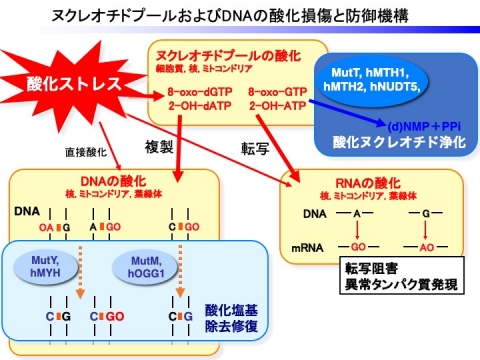
高等植物のシロイヌナズナでは、28種類存在するNudix hydrolae (AtNUDX)の中で、細胞質局在型のAtNUDX1のみが、8-oxo-dGTPに対して高い活性を示し、ヌクレオチドプール中に蓄積する8-oxo-dGTPを加水分解することで、DNAの酸化損傷を抑制していることが明らかになりました。
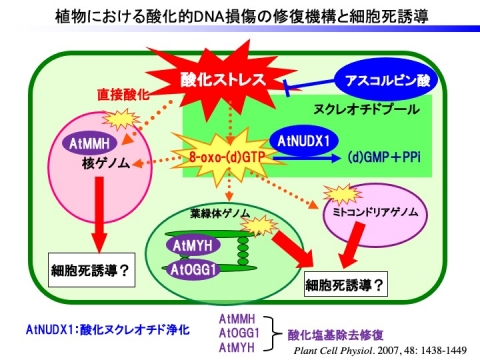
これらの研究を通して、植物の8-oxo-dGTP加水分解酵素による酸化ヌクレオチド浄化機構は、動物と比較して質的量的に劣っていることがわかりました。すなわち、動物にはMTH1をはじめ、複数の酸化ヌクレオチド加水分解酵素が、細胞質だけでなく、ゲノムの存在場所である核やミトコンドリアに複数存在ますが、シロイヌナズナではAtNUDX1が細胞質に局在するのみです。また、動物では核やミトコンドリアゲノムへの酸化損傷の蓄積は細胞死誘導の引き金になっていることから、それらゲノムの酸化障害は厳密に制御されています。
そこで、ヒトのMTH1をシロイヌナズナの葉緑体やミトコンドリアで強制発現させた結果、酸化ストレス耐性能が向上することが明らかになりました。この結果は、植物オルガネラにおけるゲノムの酸化損傷は、動物と類似して細胞死誘導の引き金になっている可能性を示すものです。
現在、AtNUDX1だけでなく、アスコルビン酸などの抗酸化物質が植物のDNA突然変異の抑制に果たす役割、および各細胞内小器官における酸化DNA修復能が植物の環境ストレス耐性に果たす役割について解析を進めています。