有用物質分解酵素、Nudix hidrolaseの生理機能
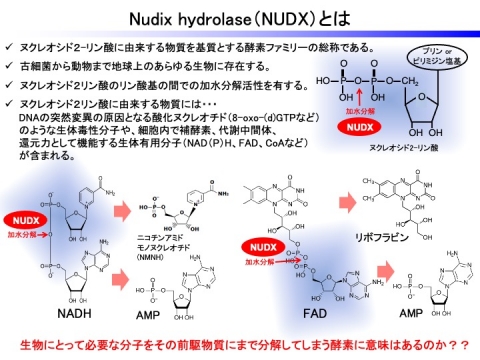
Nudix(nucleoside diphosphate linked some moiety X)hydrolaseは、ヌクレオシド二リン酸類縁体(NDP‑X)からNMP+P‑Xへの加水分解(ピロホスホハイドロラーゼ)活性を持つタンパク質ファミリーの総称であり、ウイルスからヒトにいたる300種以上の生物に、2,500を超える遺伝子が存在します。本酵素ファミリーの基質は、酸化ヌクレオチド、ADP-リボース、mRNA 5’-キャップ構造に加え、ナイアシン、パントテン酸およびフラビンなどビタミンの補酵素型であるNAD(P)H、FAD、CoAなど非常に多岐にわたります。さらに最近、葉酸生合成の代謝中間体であるジヒドロネオプテリン-3リン酸やビタミンB1の補酵素型であるチアミンピロリン酸などの非ヌクレオシド二リン酸由来の物質も基質とすることが報告されています。これらの化合物の多くは還元力/レドックス緩衝剤、シグナル分子、もしくは代謝中間体や補酵素などの役割を果たす重要な生体分子です。
生命活動を支える代謝は、生合成系と分解系の絶妙なバランスの上で成り立っています。したがって、生体内における本酵素ファミリーによる加水分解を介したさまざまな代謝や細胞応答への関与は、生命活動を支える重要な生体分子の代謝制御における分解過程の重要性を示唆するものです。さらに、各生物種におけるNudix hydrolase相同遺伝子の保有数は、Escherichia coli:13種類、Bacillus cereus: 26種類、Saccharomyces cerevisiae:7種類、Drosophila melanogaster:20種類、Homo sapiens:24種類であり、それらの有する本酵素ファミリーの総数の増加は、進化過程において当該生物が獲得した代謝能力や適応力を反映していると考えられています。しかし、動植物や微生物を含めた全ての生物において、酸化ヌクレオチドを基質とするNudix hydrolaseサブファミリーがDNA突然変異の抑制に機能していることを除いて、ほとんどのサブファミリーの生理機能は全く明らかにされておらず、推測の域を出ていないのが現状でした。
そこで我々は、植物Nudix hydrolaseファミリーの生理機能の解析を進めており、それらが種々の生体分子の分解を介した単なる代謝制御のみならず、多様な細胞応答と深く関連していることを明らかにしてきました。それらの成果は、生体有用分子の代謝制御が生合成経路だけでなく、Nudx hydrolaseによる分解経路との絶妙なバランスの上で成り立って、ホメオスタシスを維持していることを強く示すものでした。さらに、植物は生存戦略として、Nudix hydrolaseによる種々の生体分子の分解経路を巧みに発達させてきたと考えられます。今後、さらなる植物Nudix hydrolaseファミリーの機能解析を進め、それらの生物界における分布を明らかにすることで、生物の未知なる巧妙な代謝経路や生存戦略の理解の進展につながることが期待されます。
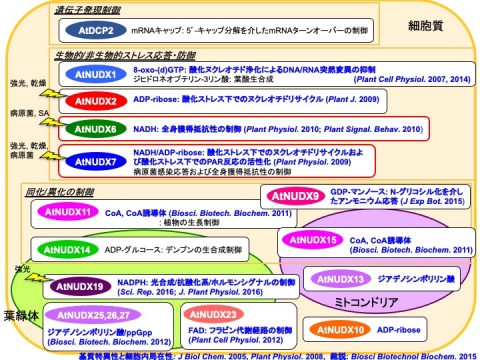
この研究の過程で、シロイヌナズナNudix hydrolaseの生理機能解析から、AtNUDX6および7はどちらもNADHを生理的基質とするにも関わらず、生体内で異なる生理機能を有することが明らかになりました。また、他の研究グループにより、AtNUDX7が病原菌感染時における植物ホルモンのサリチル酸蓄積の抑制に機能すると報告されています。また、AtNUDX19はNADPHの代謝制御を介して、光合成とストレス応答・防御系と植物ホルモンシグナリング経路のバランス制御に機能していることも明らかになりました。
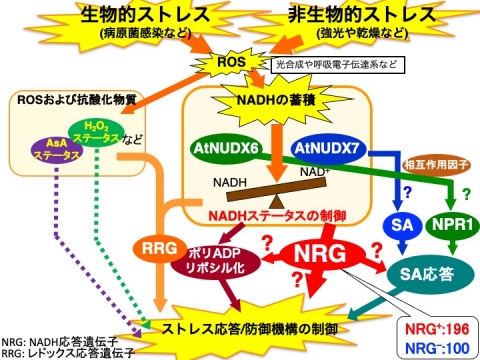
これらの事実から、各オルガネラでのNAD(P)Hの分解系による細胞内レドックスバランス制御の重要性は明確であり、特に、AtNUDX6および7による細胞質でのNADH代謝は、AtNUDX19による葉緑体内でのNADPH代謝を始めとする細胞内レドックス制御システムにより発信されたレドックスシグナルを統括する「インテグレーター」としての役割を果たしている可能性があります。
そこで、細胞内NADHレベルが段階的に増加しているAtNUDX6および7のシングルおよびダブル遺伝子破壊株を用いたトランスクリプトーム解析を行った結果、細胞内NADHレベルと発現レベルが正もしくは負に相関している遺伝子(NADH-responsive gene, NRG)を多数同定しました。現在、それらNRGの生理機能解析やNADHと既報の抗酸化物質によるレドックス制御機構の関係について解析を進めています。